仅今天一天,Nature最新一期更新了7篇中国学者的论文(第一作者或通讯作者),研究领域各点开花,中国学者迎来大爆发!其中有最强夫妻教授(麦健辉、单杰教授)年内第二篇Nature!西北大学新发现的化石将苔藓动物的地质历史前推5000万年!加州大学河边分校杨贞标、福建农林大学林文伟揭示植物细胞“酸性生长”的核心机制!吉林大学崔银秋团队在青铜时代塔里木盆地木乃伊的基因组起源取得重大突破!加州大学旧金山分校程亦凡教授团队成功探究Na+通量如何为脂质修饰的刺猬释放提供动力!中国留学生Ying Wang一作: 哺乳动物细胞中线粒体谷胱甘肽输入的分子机制!中国留学生李兰馨一作: 根生长中H+通量的细胞表面和细胞内生长素信号传导!
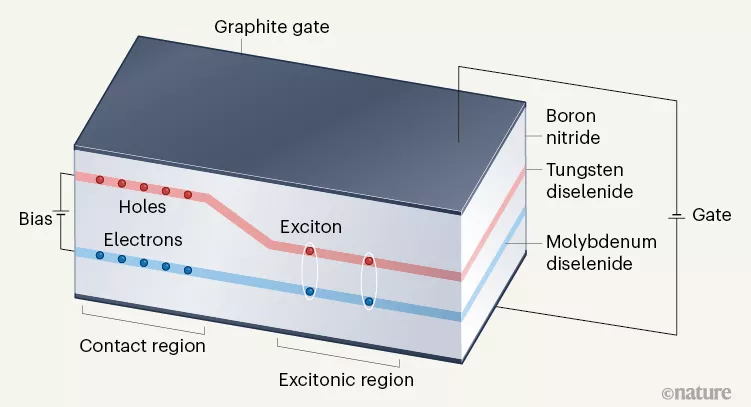
Part 1: 最强夫妻教授:原子双层中强相关激子绝缘体


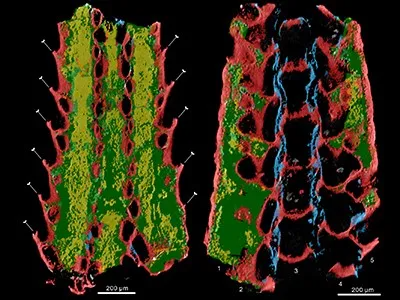
Part 2: 西北大学:化石证据揭示了苔藓虫的早期寒武纪起源

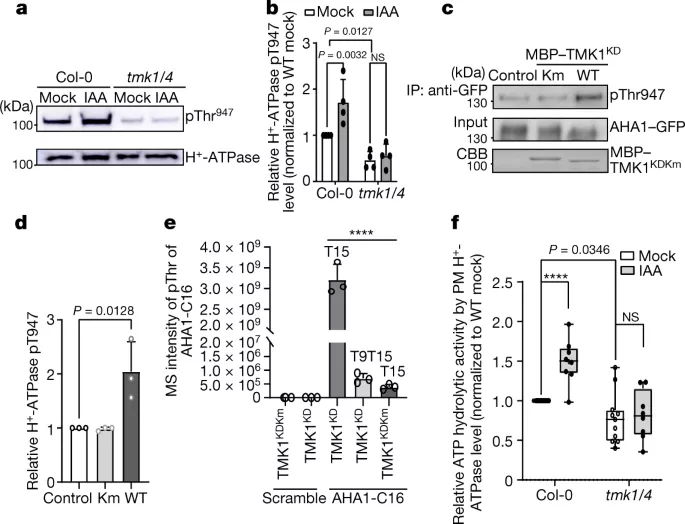
Part 3: 杨贞标团队:基于TMK的细胞表面生长素信号激活细胞壁酸化

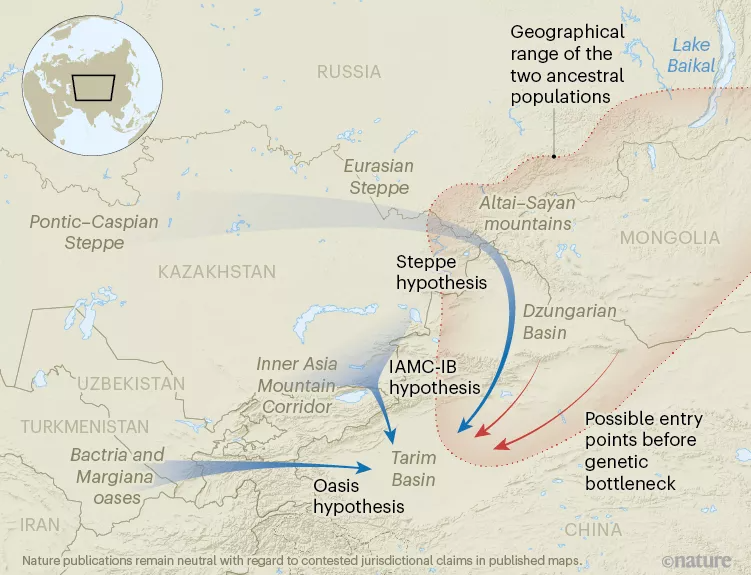
Part 4: 吉林大学崔银秋团队:青铜时代塔里木盆地木乃伊的基因组起源

在今中国西北部新疆南部的塔里木盆地偏远沙漠中发现了4000年历史的自然木乃伊个体,引发了对这些个体血统的数十年猜测。
来自吉林大学的崔银秋教授研究团队解决了木乃伊的遗传起源问题,并表明它们是古老的、遗传上孤立的种群的后裔。作者的结论对未来对亚洲内陆史前史的研究具有重要意义,这些研究必须解决文化交流与遗传祖先之间的复杂关系。研究团队展示了来自准噶尔盆地约公元前3000-2800的5个个体和来自塔里木盆地的约公元前2100-1700个个体的13个个体的基因组数据,分别代表了新疆北部和南部发现的最早的人类遗骸。研究团队发现青铜时代早期的准噶尔人主要表现出阿凡纳谢沃血统,并有额外的本地贡献,而早期-中期青铜时代的塔里木人只包含本地血统。来自小河遗址的塔里木人的牙结石中进一步显示了乳蛋白的有力证据,表明该遗址自建立以来对奶牛畜牧业的依赖。研究团队的研究结果不支持先前关于塔里木木乃伊起源的假设,这些木乃伊被认为是阿凡纳谢沃的后裔,讲原始吐火罗语的牧民,或者起源于巴克特里亚·马尔吉亚纳文明体考古群或内亚山地走廊文化。相反,尽管在青铜时代早期,阿法纳西沃移民可能将吐火罗语合理地引入准噶尔盆地,但研究团队发现塔里木盆地最早的文化似乎起源于遗传上孤立的当地人口,他们采用了邻近的牧民和农业实践,这使得他们沿着塔克拉玛干沙漠的河流绿洲定居和繁荣。
Part 5: 程亦凡团队: Na+通量如何为脂质修饰的刺猬释放提供动力

Dispatched蛋白与NPC1和PTCH1胆固醇转运蛋白以及RND家族的H+驱动转运蛋白相关,通过从胚胎表达的紧密定位位点释放脂质修饰的Hedgehog蛋白,使其具有组织模式化活性。来自加州大学旧金山分校的程亦凡教授和斯坦福大学Philip A. Beachy教授确定了小鼠蛋白质Dispatchedhomologue1(DISP1)的冷冻电子显微镜结构,揭示了三个Na+离子在穿过其跨膜结构域的通道内协调。研究团队发现Hedgehog输出的速率取决于跨质膜的Na+梯度。DISP1-NNN中的跨膜通道和Na+结合被破坏,DISP1-NNN是一种用天冬酰胺替代三个膜内天冬氨酸残基的变体,每个残基都协调并中和三个Na+离子之一的电荷。DISP1-NNN和破坏单个Na+位点的变体保留与SCUBE2受体的结合,但在将脂质修饰的Hedgehog蛋白输出到SCUBE2受体方面受到损害。声波刺猬蛋白(ShhN)的氨基末端信号域与DISP1的相互作用通过广泛的埋藏表面积和与扩展的弗林蛋白酶裂解DISP1臂的接触发生。变异性分析表明,ShhN结合仅限于一系列连续的DISP1构象中的一个极端。结合和未结合的DISP1构象显示出不同的Na+位点占有率,这表明跨膜Na+通量可以为从膜中提取脂质连接的Hedgehog信号提供动力的机制。DISP1中的Na+配位残基在PTCH1和其他后生动物RND家族成员中是保守的,这表明Na+通量为它们的构象驱动活动提供动力。
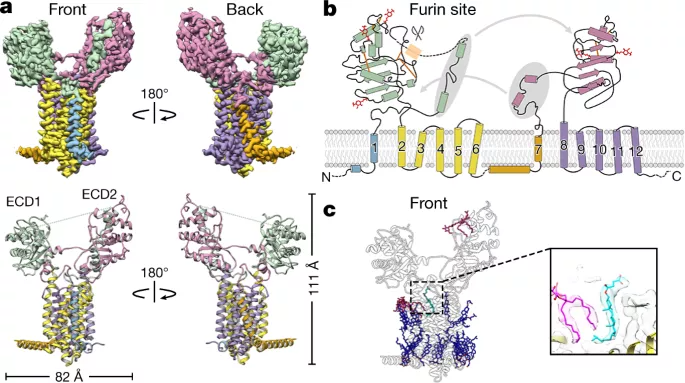
Part 6: 中国留学生Ying Wang一作: 哺乳动物细胞中线粒体谷胱甘肽输入的分子机制

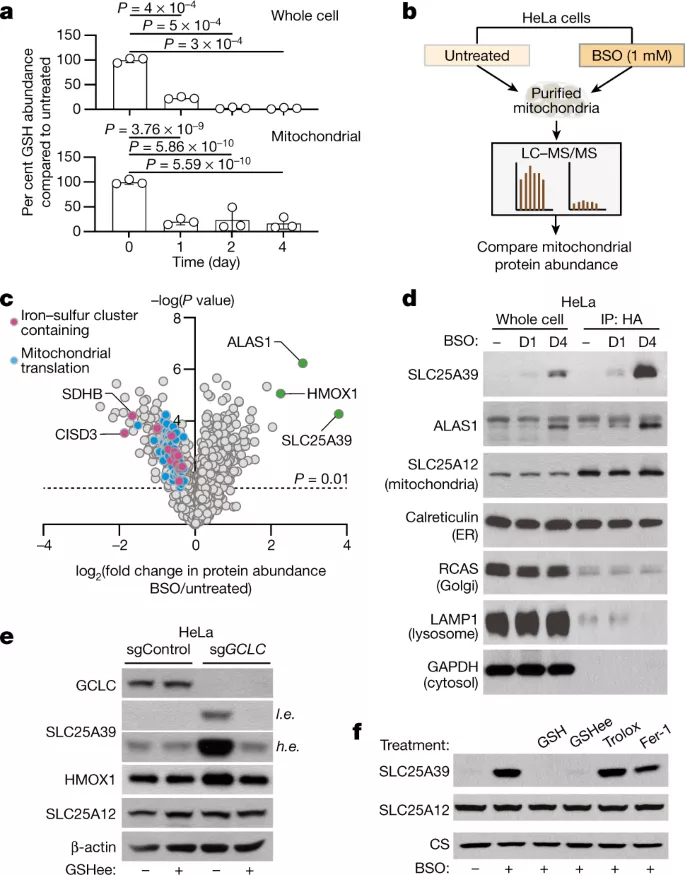
Part 7: 中国留学生李兰馨一作: 根生长中H+通量的细胞表面和细胞内生长素信号传导

生长调节使植物的发育适应其环境。一个突出的例子是对重力的反应,其中芽向上弯曲,根向下弯曲。这个悖论是基于植物激素生长素的相反作用,它通过一种未知的细胞机制促进芽中的细胞扩张,同时在根中抑制它。来自奥地利科学技术学院JiříFriml教授课题组通过在拟南芥中结合微流体、实时成像、基因工程和磷酸化蛋白质组学,研究团队加深了对生长素如何抑制根系生长的理解。研究团队表明,生长素激活两种不同的、拮抗作用的信号通路,这些通路集中在质外体pH的快速调节上,这是生长的致病决定因素。基于细胞表面的跨膜激酶1(TMK1)与质膜H+-ATPases相互作用并介导磷酸化和激活以进行质外体酸化,而细胞内的典型生长素信号促进净细胞H+流入,导致质外体碱化。这两种抵消机制的同时激活为在复杂土壤环境中进行快速、微调的生长调节奠定了基础。

1、https://www.nature.com/articles/s41586-021-03947-9
2、https://www.nature.com/articles/s41586-021-03976-4
3、https://www.nature.com/articles/s41586-021-04033-w
4、https://www.nature.com/articles/s41586-021-04052-7
5、https://www.nature.com/articles/s41586-021-04025-w
6、https://www.nature.com/articles/s41586-021-04037-6
7、https://www.nature.com/articles/s41586-021-03996-0
本文来源:高分子科学前沿


 上一篇
上一篇